

D2.1: Cell and nuclear division
Master IB Biology D2.1: Cell and nuclear division with notes created by examiners and strictly aligned with the syllabus.
IB Syllabus Requirements for Cell and nuclear division
D2.1.1 Generation of new cells in living organisms by cell division
D2.1.2 Cytokinesis as splitting of cytoplasm in a parent cell between daughter cells
D2.1.3 Equal and unequal cytokinesis
D2.1.4 Roles of mitosis and meiosis in eukaryotes
New cells come from existing cells
Cell division is a biological process in which one parent cell divides to form daughter cells. In every living organism, new cells arise when a pre-existing cell divides; in normal life, we don’t see fully functioning cells built from non-living chemicals.
A parent cell is a cell that divides to produce new cells. A daughter cell is a cell formed by division of a parent cell. You may also see the term “mother cell”, but don’t let that wording give the wrong idea: the parent cell does not stay behind as a separate individual after division. It becomes the two daughter cells.
Cell division has three main roles: growth, maintenance and reproduction. A multicellular organism grows by increasing its cell number, keeps tissues working by replacing lost or damaged cells, and may reproduce by producing cells that form offspring. In asexual reproduction, repeated mitotic divisions can produce many genetically identical cells, so a clone can behave as a single genetic individual even when it appears to have many separate parts.
Dividing the cytoplasm
Cytokinesis is the division of the cytoplasm of a parent cell between daughter cells. It differs from nuclear division: mitosis or meiosis divides nuclei; cytokinesis divides the cell body.
In animal cells, a contractile ring sits just inside the plasma membrane. Actin and myosin proteins make up the ring. As it contracts, the ring pulls the membrane inward at the cell equator and creates a cleavage furrow. Once the furrow reaches the centre, the cell pinches into two.
A rigid cell wall means plant cells can’t simply pinch apart. Instead, vesicles gather at the equator of the cell and fuse, forming new plasma membrane between the daughter cells. The vesicles also carry materials used to build the new cell wall between them. The key comparison is straightforward: animals pinch the membrane inwards; plants build a separating plate outwards across the middle.

Equal division is common, but not universal
Equal cytokinesis is cytokinesis where the cytoplasm is split into approximately equal portions between daughter cells. You see this often in tissues where the daughter cells will do similar jobs, such as cells in a growing root tip.
Unequal cytokinesis is cytokinesis where one daughter cell gets substantially more cytoplasm than the other. The smaller daughter cell can survive only if it gets a nucleus and at least one of each organelle that cannot be made from scratch. Mitochondria are the classic example: a cell makes new mitochondria by growth and division of existing mitochondria, so a daughter cell with none would be in real trouble. The same idea applies to chloroplasts in plant cells that need them.
Budding in yeast
Budding is a form of asexual reproduction where a smaller daughter cell grows out from a larger parent cell. In yeast, the nucleus divides by mitosis, one nucleus moves into the bud, and a wall forms between the bud and the larger cell. The bud receives only a small share of cytoplasm, so this is unequal cytokinesis.
Oogenesis in humans
Oogenesis is the production of egg cells in ovaries by meiotic divisions and cell differentiation. In human oogenesis, cytokinesis is strongly unequal. Most of the cytoplasm stays in one large cell that can become an oocyte, while tiny polar bodies receive nuclei but very little cytoplasm and usually do not develop further. This fits the biology: the egg cell needs reserves for early development, whereas the polar bodies provide a way to discard extra chromosome sets.

Nuclear division must come before cell division
Nuclear division is the division of a nucleus, allocating genetic material to new nuclei. If a eukaryotic cell split its cytoplasm before dividing its nucleus, one daughter cell could end up with no nucleus. An anucleate cell is a cell that lacks a nucleus. Without a nucleus, these cells cannot transcribe nuclear genes, so they cannot make the full range of proteins needed for long-term growth and maintenance.
Mitosis is nuclear division that produces genetically identical nuclei while maintaining chromosome number. Eukaryotes use it for growth, replacement of cells, tissue repair and asexual reproduction. Mitosis preserves the genome: daughter nuclei receive the same genes as the parent nucleus.
The symbol n is the haploid chromosome number, where n is the number of different chromosome types in one complete set (dimensionless count). A diploid eukaryotic body cell has 2n chromosomes. When a diploid nucleus undergoes mitosis, it produces diploid daughter nuclei, so chromosome number is maintained.
Meiosis is nuclear division that halves chromosome number and generates genetically varied nuclei. Sexual life cycles use it to produce nuclei that can contribute to gametes. Meiosis changes a diploid nucleus into haploid nuclei, so fertilization can restore the diploid number rather than doubling the chromosome number every generation.

Organisms grow through several linked processes: cells divide by mitosis, cytokinesis splits the cytoplasm, and the resulting cells grow and specialize. In plants, enlargement of cells and production of biomass also matter, but mitosis and cytokinesis provide the new cells on which those processes act.
Chromosomes must be copied first
DNA replication is the synthesis of a new DNA molecule using an existing DNA molecule as a template. Before mitosis and meiosis, all nuclear DNA is replicated, so the genetic information can be passed on to daughter nuclei.
A chromosome is a DNA molecule associated with proteins that carries a linear sequence of genes. After DNA replication, each chromosome contains two DNA molecules. A chromatid is one of the two DNA molecules in a replicated chromosome. Sister chromatids are chromatids produced by replication of the same chromosome; they are genetically identical unless a mutation has occurred.
Sister chromatids stay attached until anaphase. A cohesin is a protein complex that holds sister chromatids together after DNA replication. When cohesin is cut at the correct time, the chromatids separate and move to opposite poles.

A useful exam phrase: after replication, a chromosome has two chromatids, but it is still counted as one chromosome until the chromatids separate. Once they separate, each chromatid is considered a chromosome in its own right.
Making DNA manageable
Chromosome condensation packages long DNA molecules into shorter, thicker chromosome structures for nuclear division. This matters because DNA molecules are extremely long compared with the nucleus. Left fully extended, they could tangle, snap or fail to separate cleanly.
Histones are proteins that DNA winds around to form nucleosomes. The wrapping compacts DNA. Extra coiling and folding shorten it even more, producing visible chromosomes during nuclear division.
Moving chromosomes
Microtubules are hollow protein fibres made from tubulin that can rapidly assemble and disassemble. In mitosis and meiosis, microtubules build a spindle that reaches from the poles of the cell towards chromosomes.
A kinetochore is a protein structure on a chromosome where spindle microtubules attach. It works as a microtubule motor as well: by removing tubulin subunits, it helps shorten attached microtubules and pull chromosomes or chromatids towards poles.
The idea is straightforward. Chromosomes condense first, so they can be moved safely. Then microtubules attach and pull. In mitosis and meiosis II, sister chromatids separate. In meiosis I, homologous chromosomes separate.

The four phases
Mitosis is usually split into four phases: prophase, metaphase, anaphase and telophase. Interphase is not part of mitosis, although DNA has already been replicated there before mitosis starts.
Prophase is the phase of mitosis in which chromosomes condense and the mitotic spindle begins to form. By the end of prophase, the nuclear membrane breaks down, so spindle microtubules can interact with chromosomes.
Metaphase is the phase of mitosis in which chromosomes align at the equator of the cell. Sister chromatids attach to opposite spindle poles, and tension checks that the attachment is correct. Cohesin still holds the sister chromatids together.
Anaphase is the phase of mitosis in which sister chromatids separate and move to opposite poles. Cohesin is cut, and kinetochore microtubules shorten. From this point onwards, the separated chromatids are chromosomes.
Telophase is the phase of mitosis in which chromosomes arrive at the poles, nuclear membranes reform and chromosomes decondense. Cytokinesis often starts during telophase, so by the end of mitosis the cell is usually already well on the way to becoming two cells.

The process produces two genetically identical daughter nuclei because DNA was copied before mitosis and one copy of each chromosome moves to each pole.
What to look for in diagrams and micrographs
When you identify mitotic phases, look first at what the chromosomes are doing. Cell shape on its own can be misleading.
- Interphase: nucleus visible; chromosomes not individually distinct.
- Prophase: chromosomes visible as condensing threads or rods; not aligned on the equator.
- Metaphase: chromosomes aligned across the equator.
- Anaphase: two groups of chromosomes separating towards opposite poles; often V-shaped or pulled-looking.
- Telophase: two chromosome groups at poles; nuclei reforming; cytokinesis may be visible.
Plant root tips are useful for practice because the cells often sit in files. To make a root tip squash, grow roots, fix the tissue, use acid to loosen the cell walls, stain DNA so the chromosomes show clearly, then gently squash the root tip under a coverslip before viewing it under a microscope. Take care with safety: acid and stains require eye and skin protection.
You’ll use this skill with drawn diagrams, cells seen through a microscope, and micrographs. Real cells won’t look as tidy as textbook drawings, so combine several clues: chromosome condensation, alignment, separation, and whether one or two nuclei are visible.

Haploid and diploid
A haploid nucleus is a nucleus that contains one complete set of chromosomes, with no homologous pairs. A diploid nucleus is a nucleus that contains two complete sets of chromosomes, arranged as homologous pairs.
A homologous chromosome pair is a pair of chromosomes with the same genes in the same sequence, though the alleles may differ. One homologous chromosome may have come from the maternal parent and one from the paternal parent.
A reduction division is a nuclear division that reduces chromosome number by half. Meiosis is a reduction division because one diploid nucleus ultimately gives four haploid nuclei.
Why meiosis is needed
In a sexual life cycle, fertilization fuses two haploid gametes to form a diploid zygote. If gametes were diploid, chromosome number would double every generation. Meiosis prevents this by producing haploid nuclei before gamete formation or at another stage in the sexual life cycle.
Two rounds of segregation
Segregation is the separation of chromosomes or chromatids to opposite poles during nuclear division. Meiosis has two rounds of segregation:
- In meiosis I, homologous chromosomes separate. This changes a diploid nucleus into haploid nuclei because each new nucleus receives only one chromosome from each homologous pair.
- In meiosis II, sister chromatids separate. This resembles mitosis, but it occurs in haploid cells and produces four haploid nuclei in total.
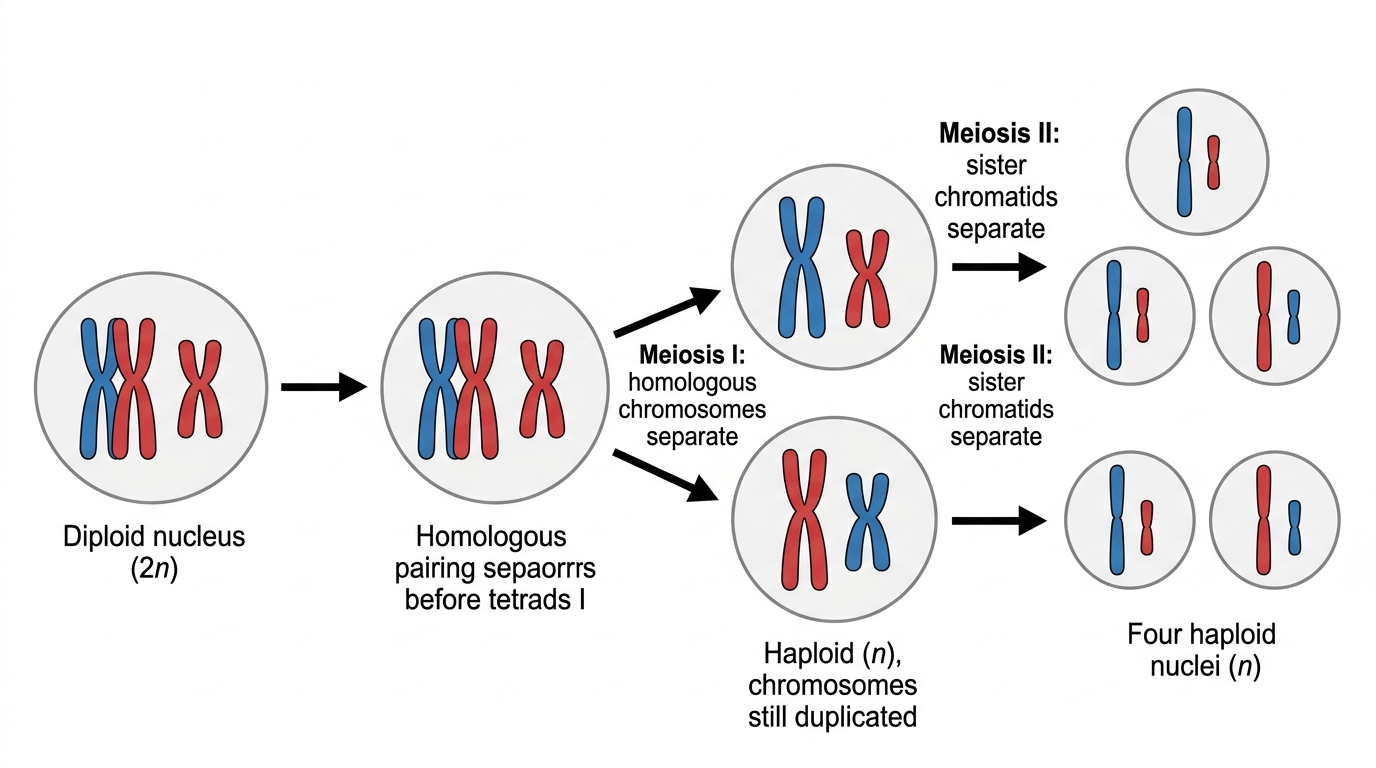

When chromosomes fail to separate
Non-disjunction is an error in nuclear division where homologous chromosomes or sister chromatids do not separate correctly. During meiosis I, a homologous pair may go to the same pole. During meiosis II, sister chromatids may go to the same pole.
This produces gametes with an abnormal chromosome number: either one chromosome extra or one chromosome missing. A missing chromosome is usually lethal because too many essential genes are absent. An extra chromosome is often lethal as well, though some cases survive.
Down syndrome is a human genetic condition usually caused by having three copies of chromosome 21. This is called trisomy 21. It can happen when a gamete carrying an extra chromosome 21 fuses with a normal gamete. Individuals vary, but possible features include effects on learning, hearing, vision and heart development.

Non-disjunction can affect sex chromosomes too, producing combinations such as XXY or a single X chromosome. For this topic, the key is not memorising every condition. It’s understanding the meiotic error: chromosomes did not segregate correctly.
Meiosis reshuffles alleles
Meiosis produces genetic diversity by making new combinations of alleles. In this section, the two key processes are crossing over and random orientation of bivalents.
A bivalent is a pair of homologous chromosomes held together during meiosis I. Synapsis is the pairing of homologous chromosomes to form bivalents. By this stage, each chromosome has already replicated, so a bivalent contains four chromatids.
Crossing over is the exchange of corresponding DNA segments between non-sister chromatids of homologous chromosomes. It happens during prophase I. At the points where chromatids cross and stay attached, a chiasma can be seen; a chiasma is a physical connection between homologous chromatids produced by crossing over.
Crossing over produces recombinant chromatids, which have allele combinations not found in either original homologous chromosome. Because crossing over can happen at different positions along chromosomes, it creates a lot of variation.

Random orientation of bivalents is the chance alignment of each homologous pair relative to the poles during metaphase I. In each bivalent, either homologous chromosome can face either pole. One bivalent’s orientation does not affect the orientation of the others.
The number of possible chromosome combinations from random orientation is 2^n, where n is the haploid chromosome number, already defined as the number of chromosome types in one complete set. In humans, n = 23, so random orientation alone can produce over eight million chromosome combinations before crossing over is even considered.

This is one way sexual reproduction supplies variation for evolution. Meiosis creates varied gametes; fertilization combines gametes; natural selection can then act on the resulting variation in phenotypes. Variation does not guarantee evolution in a particular direction, but without heritable variation there is little for selection to sort.
Increasing cell number
Cell proliferation means an increase in cell number when cell division happens faster than cell loss. In multicellular eukaryotes, proliferation relies on repeated mitosis followed by cytokinesis, so the genome is copied and distributed before the cells separate.
Growth
In early animal embryos, repeated cell division builds the embryo from a single zygote into many cells. These early divisions stand out because they raise cell number quickly and produce cells that can later specialize.
In plants, growth is focused in meristems, regions of undifferentiated plant tissue where cells keep dividing. Root and shoot apical meristems make new cells that lengthen roots and shoots. Some daughter cells stay in the meristem and continue dividing; others leave, enlarge and differentiate.

Replacement and repair
Routine cell replacement occurs in tissues where cells are continually lost. Skin gives a clear example: deeper layers produce cells, while older cells are eventually shed from the surface.
Tissue repair also needs cell division. During wound healing in skin, surviving cells close to the wound divide to replace cells that were lost. If the damage is too severe and the relevant dividing cells are lost, repair becomes much harder.
So, to answer the broader growth question: organisms grow through coordinated cell proliferation, cell enlargement, biosynthesis of cell materials and differentiation. Cell division provides more cells; growth processes then give those cells size and function.
The repeating cycle used for proliferation
The cell cycle is the ordered sequence of events in which a cell grows, replicates its DNA, divides its nucleus and then divides its cytoplasm. Cell proliferation happens through repeated turns of this cycle.
Interphase is the period of the cell cycle between one mitosis and the next. It includes G1, S and G2.
G1 phase is the stage of interphase after mitosis and before DNA replication. During this stage, the cell grows and carries out normal metabolism. Each chromosome consists of one DNA molecule.
S phase is the stage of interphase during which nuclear DNA is replicated. After S phase, each chromosome consists of two sister chromatids held together.
G2 phase is the stage of interphase after DNA replication and before mitosis, when the cell continues preparation for nuclear division.
After G2, mitosis occurs, followed by cytokinesis. Some cells leave the repeating cycle and enter G0, a non-dividing state in which they may differentiate and carry out a specialized function.

Data on DNA content per nucleus can be used to infer cell-cycle stage. Cells in G1 have the lower DNA content. Cells in G2 and early mitosis have about double that DNA content, while cells in S phase fall between these values because replication is still in progress.
Interphase is busy, not resting
Interphase is metabolically active. The cell may not be visibly dividing, but it is still doing the work needed for division later on.
During interphase, growth depends on biosynthesis, the enzyme-controlled production of cellular molecules from simpler substances. The cell makes proteins, including enzymes and structural proteins. During S phase, it also makes DNA, as well as membrane components such as phospholipids and membrane proteins.
The cytoplasm increases in volume, and the cell must increase organelle numbers before it divides. Mitochondria and chloroplasts matter here because they are produced by growth and division of pre-existing mitochondria and chloroplasts. A cell that has none cannot simply build them from loose parts.
Chromatin state matters too. Chromatin is DNA associated with proteins in a less condensed form within the nucleus. When chromatin is decondensed, it can be transcribed, allowing protein synthesis. Some DNA stays more condensed as heterochromatin where genes are not needed for that cell’s current activities.

Cyclins rise and fall
Cyclins are regulatory proteins that rise and fall in concentration during the cell cycle, helping control movement through checkpoints. The syllabus doesn’t ask for the detailed roles of named cyclins, so keep the idea at the right level: different cyclins peak at different times.
A checkpoint is a control point in the cell cycle where progression is allowed only when specific conditions have been met. A cell passes a checkpoint once the relevant cyclin reaches a threshold concentration. If that threshold is not reached, the cell stays in its current stage.
This control matters because cell division is powerful. Too little proliferation prevents growth and repair; too much proliferation risks tumour formation. Cyclins help keep the sequence orderly: growth, DNA replication, preparation for mitosis, mitosis and cytokinesis.

When control genes mutate
A mutation is a change in the base sequence of DNA. When mutations happen in genes that control the cell cycle, the timing of division, and when it stops, can be disrupted. A cell that loses control may keep dividing, and its daughter cells inherit the same faulty control.
A proto-oncogene is a normal gene that promotes cell division or cell-cycle progression when appropriately regulated. An oncogene is a mutated or overactive form of a proto-oncogene that stimulates excessive cell division. One overactive copy can be enough to increase risk, because the faulty signal actively pushes the cell cycle forward.
A tumour suppressor gene is a gene whose normal product inhibits cell division, supports DNA repair or promotes death of badly damaged cells. When tumour suppressor genes lose function, the “brakes” on division are weakened. Usually both copies must be affected before control is seriously lost, because one working copy may still produce enough functional protein.
Mutagens such as some chemicals and high-energy radiation increase mutation rates, so there is a higher chance that cell-cycle control genes are damaged. Tumour formation usually requires several mutations to accumulate in the same cell lineage, not just one unlucky change. Every division of a partly deregulated cell makes more cells in which further mutations can occur.
Tumours are not all the same
A tumour is a mass of cells formed by uncontrolled cell division. Tumours vary: some have cells that divide quickly, some grow more slowly, and some stay as a single mass while others spread.
A primary tumour is the original tumour at the site where uncontrolled division first began. A benign tumour is a tumour that remains localized and does not invade neighbouring tissues or spread to distant sites. Benign tumours are not classed as cancer, although they can still cause problems if they press on tissues.
A malignant tumour is a tumour whose cells can invade nearby tissues and spread to other parts of the body. Metastasis is the spread of tumour cells from one part of the body to another. A secondary tumour is a tumour that forms after metastatic cells settle and divide at a new site. Malignant tumours are cancers.

Mitotic index
The mitotic index is the ratio of cells in mitosis to the total number of observed cells in a sample. It is calculated as:
mitotic index = number of cells in mitosis / total number of observed cells
To find it, use a micrograph or microscope field, count the total number of cells, then count how many are in mitosis and divide. In practice, this could be done using a root tip meristem or tumour micrograph. A higher mitotic index shows that a larger fraction of cells are actively dividing at that moment, which often suggests more rapid proliferation.

Cancer treatments such as chemotherapy often target mitosis because cancer cells divide frequently. The problem is that normal rapidly dividing cells can be damaged too, which explains many side effects.
Were those notes helpful?
The Fast Track To Your
Best IB Coursework & College Essays
All content on this website has been developed independently from and is not endorsed by the International Baccalaureate Organization. International Baccalaureate and IB are registered trademarks owned by the International Baccalaureate Organization.